Cyanobacteria - colloquially also called blue-green algae - can produce oil from water and carbon dioxide with the help of light. This is shown by a recent study. The result is unexpected: Until now, it was believed that this ability…
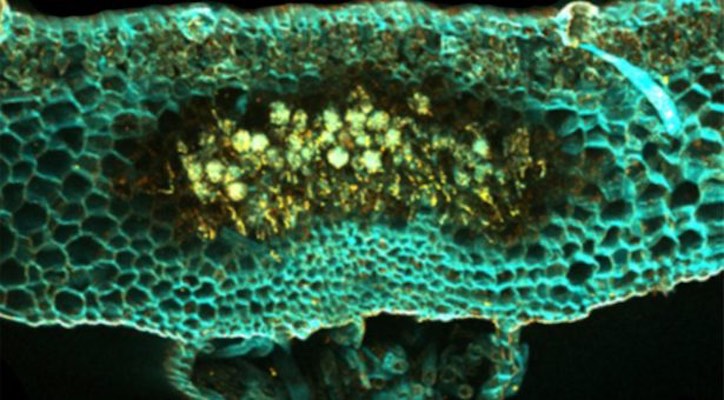
For billions of years life on Earth was restricted to aquatic environments, the oceans, seas, rivers and lakes. Then 450 million years ago the first plants colonised land, evolving in the process multiple types of beneficial relationships with microbes in…
How do plants sense their environment? How do they cope with different challenges, like competition for resources or pest attacks? In a new study researchers explain the role of ST2a, a gene up-regulated by plant proximity cues, which participates in…
Root hairs are tubular polarized extensions of root epidermal cells and are crucial for plant anchorage, nutrient acquisition, and environmental interactions. The plant hormone jasmonate has been reported to promote root hair growth. However, it remains unclear about the molecular…
New research provides a better understanding of how chemicals thought to impart unique health benefits to plants in the cabbage family are broken down to promote growth in conditions lacking sufficient sulfur. This findings could aid in the future development…
Proteins play a key role in controlling all aspects of life including plants. A team of scientists has now mapped around 18,000 of all the proteins found in the model plant Arabidopsis thaliana
Astronauts in space generally live on processed, pre-packaged space rations such as fruits, nuts, chocolate, shrimp cocktails, peanut butter, chicken, and beef to name a few. These have often been sterilized by heating, freeze drying, or irradiation to make them…
Tropical forest trees are the centerpiece of debates on conservation, climate change and carbon sequestration today. While their ecological importance has never been doubted, what has often been ignored is their ability to store cultural heritage. Using recent advances in…
Drifting algae in the Austral Ocean can bring invasive species to the Antarctic coasts, according to a new study. The report describes the first scientific evidence of a potentially invasive and colonial species –the marine bryozoan Membranipora membranacea- which reaches the…
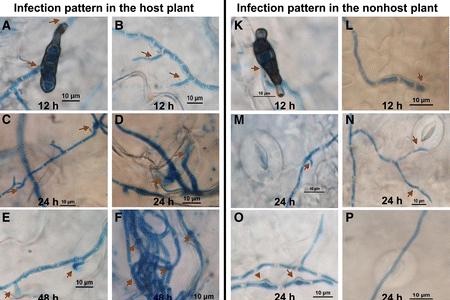
Alternaria blight caused by fungal pathogen devastates Brassica crops such as cabbage, cauliflower, broccoli, and mustard seed. Highly infectious, this fungus can infect the host plant at all stages of growth. Currently Alternaria blight is managed by chemical fungicides, but…