Botanists have long held a fascination for heterotrophic plants, not only because they contradict the notion that autotrophy (photosynthesis) is synonymous with plants, but also because such plants are typically rare and ephemeral. However, it is still a matter of debate as to how these plants obtain nutrition.
A research team consisting of Kobe University’s Associate Professor SUETSUGU Kenji (of the Graduate School of Science’s Department of Biology), Research Fellow MATSUBAYASHI Jun (of Japan Society for the Promotion of Science) and Professor TAYASU Ichiro (of the Research Institute for Humanity and Nature), has investigated the carbon age (the time since the carbon was fixed from atmospheric CO2by photosynthesis) in some non-photosynthetic mycoheterotrophic plants. Many orchids have lost photosynthetic ability and evolved an enigmatic mycoheterotrophic lifecycle. Mycoheterotrophic plants usually obtain carbon from other photosynthetic plants through a shared mycorrhizal fungal network, while some mycoheterotrophs are believed to obtain carbon from decaying litter or dead wood by parasitizing saprotrophic fungi. However, traditional approaches have provided only indirect evidence of such nutrient transportation from dead organic matter to plants.
The current study examined the utility of radiocarbon measurements to distinguish the fungal exploitation pattern of mycoheterotrophs. Mycoheterotrophic species exploiting ectomycorrhizal fungi should take up recently synthesized photosynthates, while mycoheterotrophic plants dependent on saprotrophic fungi must obtain carbon from older sources, i.e., dead wood. Therefore, the research team calculated the carbon age of mycoheterotrophic plants, using the radiocarbon emitted from atmospheric nuclear bomb tests carried out in the 1950s and 1960s as a tracer.
Through this methodology, they revealed that the carbon in some mycoheterotrophic orchids dated from over ten years prior to the sampling period. This indicates that these orchids are relying on the 14C-enriched bomb carbon from dead wood via saprotrophic fungi. Therefore, they concluded that mycoheterotrophic plants can exploit both mycorrhizal and saprotrophic fungi, which are essential components for terrestrial ecosystems. In addition, even though the term “mycoheterotroph” has replaced the formerly misapplied term “saprophyte,” some mycoheterotrophic plants are indirectly saprotrophic! The finding overturns the traditional view and opens a new perspective for understanding how these intriguing plants have become ecologically and evolutionarily successful.
Research Background
Mutualism, or mutually beneficial interactions between species, is a ubiquitous phenomenon in all ecological systems, and almost all organisms on Earth are involved in at least one mutualistic partnership. Most terrestrial plants, from bryophytes to angiosperms, form mutualisms (interspecific cooperative interactions) with fungi, whereby the plant provides a carbon source in exchange for essential mineral nutrients (Figure 1). These mutually beneficial plant-fungi associations are called mycorrhizal mutualisms.
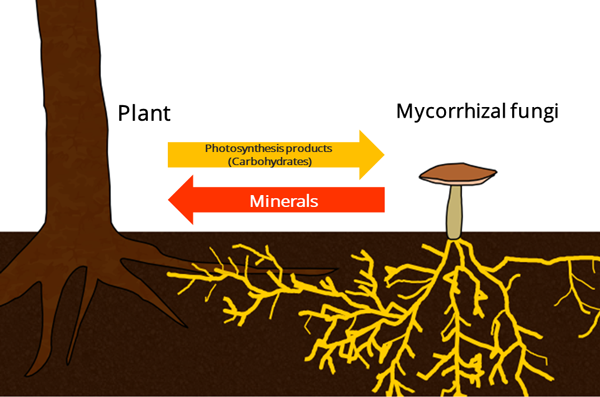
However, organisms that were originally engaged in mutualisms can sometimes turn into parasites, obtaining the benefits while delivering none in return. In mycorrhizal mutualisms, non-photosynthetic mycorrhizal plants (i.e. mycoheterotrophic plants) are considered as such cheaters since they cannot provide photosynthates to their fungal partners. Indeed, many mycoheterotrophic plants are known to obtain carbon from other photosynthetic plants through a shared mycorrhizal fungal network.
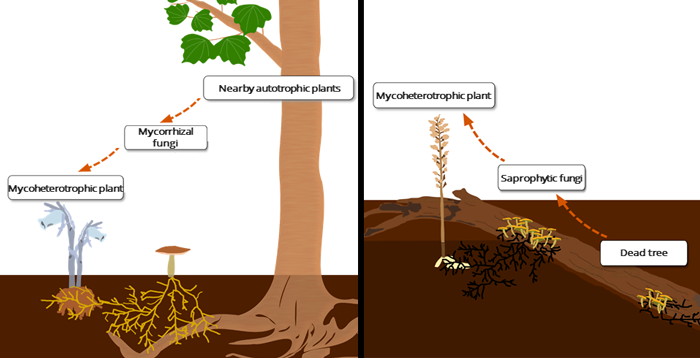
Therefore, despite their achlorophyllous nature, mycoheterotrophic plants are not directly parasitic on other plants, nor do they directly obtain carbon from rotting plant and animal matter, as was once believed (Figure 2). Nonetheless, it is known that fungi play essential roles in terrestrial ecosystems, notably as saprotrophic fungi, which decompose dead wood and decaying litter. Do some mycoheterotrophic plants dependent on these saprotrophic fungi exist? The current research team set out to illuminate this question, utilizing radiocarbon analysis.

Research Details
Radiocarbon (14C) levels could be useful for precisely estimating the trophic strategies of the symbionts of mycoheterotrophic plants, by providing a direct estimation of the mean carbon age in the biomass. Atmospheric nuclear bomb testing during the mid-20th century increased the 14C concentration in the atmosphere worldwide, the peak of which was around 1963. Then the atmospheric 14C concentration gradually decreased after the ban on atmospheric nuclear testing in 1963.
Therefore, given that the 14C content of organic matter synthesized by primary producers is the same as the corresponding 14C content of atmospheric CO2, the carbon age (the time since carbon was fixed from atmospheric CO2 by photosynthesis) can be estimated by measuring the concentration of 14C arising from the bomb tests of the 1950s and 1960s. As previously explained, mycorrhizal fungi receive photosynthesized carbon from plants. In addition, the mycoheterotrophic plants exploiting mycorrhizal fungi also obtain carbon from other nearby plants through mycorrhizal network. Therefore, the research group hypothesized that the 14C values of mycoheterotrophs exploiting mycorrhizal fungi would resemble the 14C values of atmospheric CO2 for the surrounding autotrophic (photosynthesizing) plants, since this carbon should be very recent. On the other hand, the carbon of mycoheterotrophs exploiting saprotrophic fungi (particularly wood-decaying ones) should be older and therefore contain a higher concentration of the 14C that was generated by nuclear tests.
To investigate these hypotheses, the 14C concentrations of 10 species of mycoheterotrophic plants collected across 10 sites in Japan were measured. Of these ten species, six species (two ericaceous species Monotropastrum humile and Pyrola subaphylla and four orchidaceous species Cephalanthera subaphylla, Chamaegastrodia shikokiana, Neottia nidus-avis and Lecanorchis nigricans) showed low 14C concentrations which were similar to the results for autotrophic plants- confirming that they utilize very recent carbon.
On the other hand, it was revealed that the other four species, which were all types of orchid (Gastrodia elata, Cyrtosia septentrionalis, Yoania japonica and Eulophia zollingeri), contained very high concentrations of 14C, dating from over ten years prior to the sampling period. These research results indicate that some mycoheterotrophs have acquired 14C-enriched bomb carbon from dead wood via saprotrophic fungi. This indicates that some mycoheterotrophic plants do not obtain their carbon by tapping into existing mycorrhizal networks, but recruit saprotrophic fungi into novel mycorrhizal symbioses (Figure 3).
Conclusion
This research illuminated that mycoheterotrophic plants can exploit both mycorrhizal and saprotrophic fungi, which are essential components for terrestrial ecosystems. Many botanists rejected the use of the term ‘saprophyte’ as incorrect and called the plants mycoheterotrophic plants to reflect their unique nutritional dependence on fungal carbon. In fact, there are no saprophytes that directly feed on dead organic matter. However, the radiocarbon approach provides conclusive evidence that some mycoheterotrophic orchids are indirectly saprophytic and are dependent on wood debris in the forest carbon cycle.
Read the paper: New Phytologyst
Article source: Kobe University