
Author: Liang Fang is a PhD candidate based at Wageningen University & Research (WUR), Wageningen, Netherlands. You may follow him in Twitter as @LiangFANGG and WUR as @CSA_WUR.
Respiration in plants is a process that consumes sugars (e.g., glucose) and oxygen, produces carbon dioxide (CO2) and water, and releases energy to maintain the primary metabolic and physiological functions during growth. It determines the net carbon gain for plants and the carbon efflux for whole ecosystems.
Unlike leaf respiration in the dark (Rdk), leaf day respiration (CO2 release by processes other than photorespiration; Rd) occurs simultaneously with other physiological processes, such as photosynthesis, photorespiration, reassimilation, and photoinhibition, in the daytime. For this reason, how to quantify Rd has long puzzled plant scientists. Techniques to quantify Rd have been implemented for decades, however, there is still debate about which is the best one. Direct measurements of Rd are usually conducted in the lab and require sophisticated devices. Indirect estimation of Rd is mainly based on gas exchange measurements by a portable photosynthetic system (e.g., Li-Cor 6800) and often applied to ecophysiological studies under field conditions.
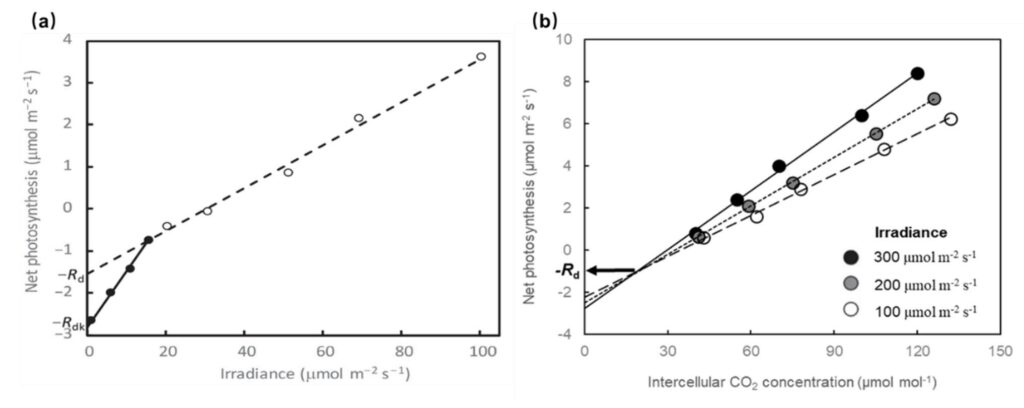
Among the methods for indirect estimation of Rd, the two best known methods are the Kok method (Kok, 1948) and the Laisk method (Laisk, 1977). The Kok method exploits the Kok effect, the abrupt break in the light response curve of net photosynthesis rate at around the light compensation point. The abrupt break is interpreted as the consequence of light inhibition to leaf respiration, and thus Rd can be calculated as the intercept of this linear relationship using points above the breakpoint, while the linear relationship below the breakpoint is interpreted as Rdk (Figure 1a; Yin et al., 2020). In the Laisk method, the response of net photosynthesis rate to low intercellular CO2 concentration is obtained at several (commonly three) levels of irradiances. These curves theoretically intersect at a common point, where the value of the net photosynthesis rate represents Rd (Figure 1b).
The Rd estimate by the Kok method is often somewhat lower than the estimate by the Laisk method (Yin et al., 2011), probably because the Kok method assumes that the PSII electron transport efficiency is constant across light levels. Then a modified Kok method, now known as the Yin method, was developed to overcome this weakness of the Kok method by incorporating the information from chlorophyll fluorescence measurements that account for the decline of the PSII electron transport efficiency with increasing light intensity. The Yin method gives Rd estimates like those the Laisk method gives (Yin et al. 2011), with the benefit that the measurements are easier and less time-consuming to implement than those with the Laisk method (Yin et al., 2011). However, theoretically and strictly speaking, both the Kok and the Yin methods can only be applied for non-photorespiratory conditions because they do not account for reassimilation of the respired CO2 and, therefore, would underestimate real Rd under common photorespiratory conditions.
Recently, Fang et al. (2022) introduced a nonrectangular hyperbolic fitting method (the NRH method; see Yin & Struik 2009) to estimate Rd under common photorespiratory conditions. The NRH method is based on a coupled biochemical photosynthesis and mesophyll conductance model, which can account for reassimilation (von Caemmerer, 2000; Yin et al., 2021). The NRH method only requires gas exchange and chlorophyll fluorescence data, which are easily accessible, even under field conditions. By using the NRH method, Fang et al. (2022) demonstrated that when the Yin method is wrongly applied to photorespiratory conditions, Rd would be underestimated by 20%, while the Kok method always underestimates Rd by 34% and 18% for photorespiratory and non-photorespiratory conditions, respectively. Besides, Fang et al. (2022) assessed the extent to which Rd of wheat leaves acclimated to drought and growth temperature, and found that drought exerted a greater influence than growth temperature on the temperature response of Rd. The findings provide information for the modelling of Rd, and thus of carbon cycling in (agro)ecosystems, under future climate change.
References
- von Caemmerer, S. (2000) Biochemical models of leaf photosynthesis. Collingwood: CSIRO Publishing.
- Fang, L., Yin, X., van der Putten, P.E.L., Martre, P. & Struik, P.C. (2022) Drought exerts a greater influence than growth temperature on the temperature response of leaf day respiration in wheat (Triticum aestivum). Plant, Cell & Environment, in press. https://doi.org/10.1111/pce.14324
- Kok, B. (1948) A critical consideration of the quantum yield of Chlorella‐ photosynthesis. Proefschrift ter verkrijging van de graad van doctor in de wis‐ en natuurkunde aan de Rijksuniversiteit te Utrecht… door Bessel Kok. The Hague: W. Junk.
- Laisk, A.K. (1977) Kinetics of photosynthesis and photorespiration in C3‐ plants [In Russian]. Moscow: Nauka.
- Yin, X., Busch, F.A., Struik, P.C. & Sharkey, T.D. (2021) Evolution of a biochemical model of steady‐state photosynthesis. Plant, Cell & Environment, 44(9), 2811–2837.
- Yin, X., Niu, Y., van der Putten, P.E.L. & Struik, P.C. (2020) The Kok effect revisited. New Phytologist, 227, 1764–1775.
- Yin, X. & Struik, P.C. (2009) Theoretical reconsiderations when estimating the mesophyll conductance to CO2 diffusion in leaves of C3 plants by analysis of combined gas exchange and chlorophyll fluorescence measurements. Plant, Cell & Environment, 32, 1513–1524. (Corrigendum in Plant, Cell & Environment, 33, 1595).
- Yin, X., Sun, Z., Struik, P.C. & Gu, J. (2011) Evaluating a new method to estimate the rate of leaf respiration in the light by analysis of combined gas exchange and chlorophyll fluorescence measurements. Journal of Experimental Botany, 62, 3489–3499.
Read the paper: Plant, Cell & Environment
Image credit: Wikimedia



